DARI SUNDA KE SAHUL
Gelombang Kedua
Sebagian besar garis keturunan populasi Gelombang Pertama sekarang tersebar luas di wilayah terdekat, seperti Jawa, Sumatra, Borneo, juga termasuk interior Semenanjung Melayu dan Thailand. Sebagian dari mereka meninggalkan Sundaland menuju arah timur, dan juga ke arah utara. Mereka meninggalkan jejakhaplotype yang berumur sangat tua, berasal dari daerah di mana haplotypetersebut ditemukan (autochthonous), dan mendukung pandangan bahwa perkembangan in situ mtDNA Australasia terjadi di sekitar Wallacea dan Sahul.
Berdasarkan keragaman genetik, garis keturunan Gelombang Pertama menyebar ke arah utara melalui Semenanjung Melayu, Thailand, Kamboja, serta Laos, dan ke arah timur, yaitu Nusa Tenggara, kepulauan Maluku, dan kemudian sampai di paparan Sahul. Berdasarkan sebaran mtDNA dan keragamannya, bahwa pusat sebaran atau asal-usul mereka adalah area di antara pulau Jawa dan Sumatra. Keragaman mtDNA populasi aborigin Kamboja menunjukkan migrasi dari arah selatan untuk mtDNA tertua di sana (M68, M69, M78, N7: Zhang et al., 2014), keragaman mtDNA populasi aborigin Malaysia juga menunjukkan hal yang sama (Baer et al., 2010). mtDNA populasi aborigin Australia adalah subclade darimacro-haplogroup N* (mtDNA O/N12 di daerah gurun Australia Tengah, mtDNA S menyebar di segala penjuru Australia sampai Tasmania, mtDNA N13 dan N14 di wilayah Kimberley) serta subclade dari macrohaplogroup M* (M42 di sekitar Teluk Carpentaria, serta M14 dan M15 di Arnhem serta Kimberley). Saya masih sulit menentukan hubungan populasi Nusa Tenggara (mtDNA N* dan M*) dengan M* dan N* yang di Jawa dan Sumatra (Bangka/Palembang). Keragaman mtDNA N* dan M* tersebut belum dikaji, kita tidak tahu apakah mereka berbagi haplotypeyang sama, atau perbandingan polimorfisme HVS-I antara ketiga area tersebut. Atau mungkin karena mereka tidak yakin secara persis sub-haplogroup apa yang akan ditujukan dalam M* dan N* tersebut. Namun yang pasti, mtDNA M dari India berbeda dengan M yang ada di daratan Asia Tenggara, serta N dari Eurasia berbeda dengan N yang ada di Australasia. Jika M dari India dan N dari Eurasia berkerabat dekat dengan L3 dari Afrika, namun tidak halnya dengan M dari Asia Tenggara dan N dari Australasia, lalu apa hubungan mereka dengan Afrika? Mengapa para ahli tidak meneliti kembali genome seluruh manusia modern untuk membuktikan bahwa garis keturunan Australia memang berbeda dari mereka yang ada di Afrika?
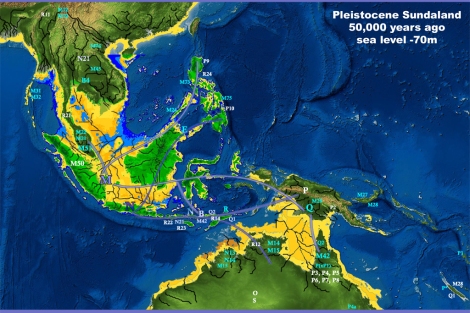
Gelombang kedua ditandai dengan migrasi ke arah timur, mengingat mereka mencapai pulau-pulau bagian timur Indonesia, New Guinea, dan juga Australia setidaknya ~50.000 tahun yang lalu. Namun migrasi ini bisa jadi tidak perlu menyeberangi laut, karena formasi kepulauan saat air laut berada > 70m di bawah permukaan saat ini, akan mempermudah mereka untuk menyeberangi Wallace’s Line melalui Borneo, Sulawesi Selatan, dan kemudian Flores, dan akhirnya mencapai Sahul (Satu eksperimental rakit bambu meniru teknologi kuno mampu mencapai Australia dari pulau Timor dalam enam hari). Bisakah mereka menyeberang ke Australia tanpa menyeberangi laut? Kemungkinan bisa jika mereka masuk melalui Papua, namun kemungkinannya agak kecil karena kekerabatan genetik yang hanya parsial antara aborigin Australia dan populasi Papua (populasi di sekitar Teluk Carpentaria; mtDNA M42 yang sepertinya berkerabat dekat dengan M29’Q karena keduanya hanya satu mutasi dari mtDNA M yang bisa jadi adalah M dari India karena ditemukannya garis keturunan M42 di sana. Sedangkan mtDNA Q di Australia hanya Q2b, dan P3b ditemukan di PNG dan Australia). Gelombang kedua ini digambarkan sedikit oleh Zhang et al. (2014) dengan sebaran mtDNA P di Sahul, Wallacea dan Filipina. Tentunya ada juga sebaran mtDNA yang lain yang kontemporer dengan sebaran mtDNA P, seperti mtDNA Q (Friedlaender et al., 2005). Untuk sebaran dan keragaman Y-DNA, Karafet et al., 2014 telah membuat pemetaan garis keturunan Y-DNA K2b1 yang migrasi dari Sunda ke Sahul.
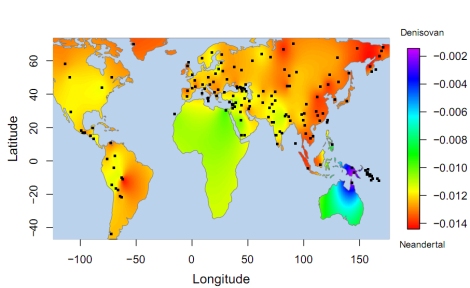
Lokasi populasi dengan materi genetik dari Denisovan
Perhatikan sebaran populasi yang mewarisi genetik dari Denisovan (Teluk Carpentaria di Australia, dan seantero Papua)
Berdasarkan penelitian Karafet et al. (2014) Y-DNA M dan S (subclade dari K2b1) bisa jadi hidup di Sundaland beberapa waktu sebelum akhirnya memutuskan untuk menyeberangi Wallace’s Line. Hal yang sama berlaku pada subclade mtDNA M (M14, M15, M29’Q, M27 dan M28, serta M42) dan N (R, B, P, O, S, N13, N14), sehingga kemungkinan besar mtDNA M dan N berasal dari Sundaland, yang mengalami difersivikasi selama perjalanan menuju Sahul, kandidat lokasi paling kuat adalah daerah antara Flores dan Alor, karena hampir semua subclademtDNA N dan M bisa ditemukan di sana. Seperti M42 di Adonara, R di Alor, dan O/N12 di Flores. Y-DNA C-RPS4Y* yang cukup beragam frekuensi dan garis keturunannya juga ditemukan di Flores, sedangkan F-P14* di Adonara dan Timor menunjukkan bahwa migrasi mereka melalui jalur selatan menuju Sahul mendapat dukungan bukti yang cukup dari penanda genetik.
Perjalanan mtDNA N (dan subclade-nya) ke arah timur sampai Australia melalui jalur selatan (Jawa, Bali, Nusa Tenggara, Timor, dan akhirnya Australia) dengan sebaran mtDNA R, B, P, O, S, N13, N14; sedangkan perjalanan mtDNA M ke arah timur melalui jalur tengah (Borneo selatan, Sulawesi selatan, Sulawesi Tengah, Maluku, kepala burung Irian, Papua Nugini serta Melanesia) dengan sebaran mtDNA Q, M27, M28, M29, sebelumnya M42, M14 dan M15 bisa jadi mendahului mereka, yang menuju Papua Nugini, langsung menuju Australia.
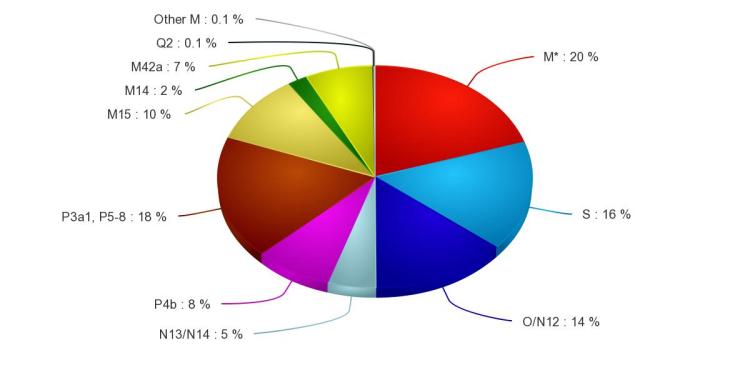
Australia mtDNA haplogroups
Seberapa dekat kekerabatan mtDNA populasi aborigin Australia denganhighlander PNG atau Papua secara keseluruhan? Mari kita mulai darimacrohaplogroup M:
M42, ditemukan di wilayah timur laut Australia, di sekitar Teluk Carpentaria, dan juga ada beberapa di populasi Adonara (Flores), namun tidak ada di PNG. Flores (NTT) bisa jadi melting pot mtDNA M karena mtDNA M42 hanya 7 mutasi langsung dari macrohaplogroup M dengan mutasi SNP 9156 pada posisi turunan. Bahkan ada cukup banyak sample populasi di NTT yang masih tidak bisa diklasifikasikan dalam macrohaplogroup M. Dengan laju mutasi di Australasia berkisar antara 6-8000 tahun, maka bisa disimpulkan mtDNA M42 berada di NTT minimal 56-58.000 tahun yang lalu. Hudjashov et al. (2007) mengestimasi umur M42a di Australia sekitar 40.600 tahun, 10 mutasi dari M42 dengan mutasi pada SNP 12771 pada posisi ancestral (yang berarti lahir di Australia), cukup sesuai dengan umur mtDNA M42 ketika pertama kali sampai di Australia. Hudjashov et al. juga mengestimasi mtDNA P berumur 55.000 tahun, N13, N14, S and O sekitar 57.000 tahun. Merriwether et al. (2005) mengestimasi umur M42 sekitar 61.700 tahun, dan M42a sekitar 54.000 tahun (lebih tua dari estimasi Hudjashov et al. (2007). van Holst Pellekaan (2010) mengestimasi M42a 39.744±9163 tahun dengan frekuensi tertinggi berada di wilayah Riverine (jauh di sebelah selatan Teluk Carpentaria). M42a terdapat di Gurnu (penutur Paakintji bagian utara), beberapa garis keturunan maternal Ngiyambaa dan mereka yang menghuni bagian barat Wiradjiri, meskipun masih perlu diverifikasi. M42 tidak berkerabat dekat secara khusus dengan M di PNG atau Melanesia dan kepulauan Pasifik, kecuali hanya mereka sama-sama garis keturunan dari macrohaplogroup M. Jalur yang ditempuh mtDNA M*(xM42) sepertinya melalui Tanimbar sebelum menginjak Sahul dan kemudian ke arah selatan melalui jembatan yang menghubungkan PNG dan Australia, dan pada akhirnya menghuni sekitar Teluk Carpentaria, di ujung utara Queensland. M42 dan M74 (M74a di China Selatan dan Vietnam) adalah keturunan dari M42’74, mereka berpisah di Sundaland, dengan M74 bergerak ke arah utara dan timur laut (M74b di Mamanwa, Filipina). Populasi aborigin Australia di sekitar Teluk Carpentaria (M42a) dan Mamanwa (M74b sama-sama mewarisi gen dari Denisovan (Kebetulan? Tentu tidak. Di India (Dravida, Austro-asiatik) ditemukan M42b, perlu kajian lebih lanjut hubungan M42b dengan garis keturunan M42 yang lain. mtDNA M42, O/N12 bersama dengan Y-DNA C-RPS4Y* (garis keturunan pria Adonara dan Flores di dominasi Y-DNA C-M38* dan C-RPS4Y*, sedangkan aborigin Australia di daerah Arnhem didominasi Y-DNA C-M374) adalah populasi tertua di Wallacea dan Sahul, merekalah yang membuat aborigin Australia (terutama Arnhem) dan populasi di NTT (Adonara dan Flores) berkerabat dekat secara genetik. Hal tersebut yang menjadikan NTT secara genetik sebagai daerah melting pot.
M14 dan M15, banyak ditemukan di bagian utara Australia sekitar Arnhem, M14 3 mutasi dari macrohaplogroup M, sedangkan M15 13 mutasi dari M. Sebagian M15 bergerak ke arah barat di sekitar Kimberley. Kedua mtDNA ini memang belum dilakukan kajian yang mendalam, namun bisa disimpulkan bahwa keduanya adalah haplogroup yang langka di Australia.
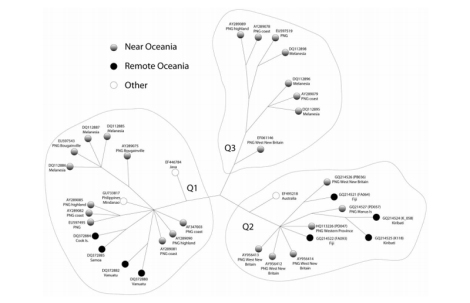
Sebaran mtDNA Q di Oceania (Corser et al., 2010)
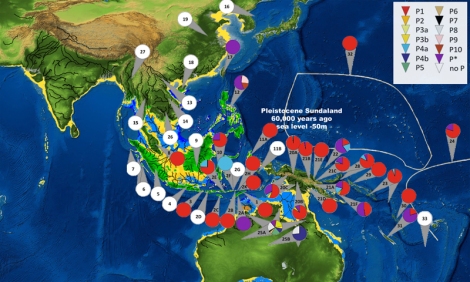
Q, mtDNA Q ditemukan tersebar di Indonesia Timur, Filipina, PNG, Melanesia dan kepulauan Pasifik, serta hanya satu subclade di Australia (Q2b di Teluk Carpentaria; Hudjashov et al. 2007). mtDNA Q hanya 3 mutasi dari macrohaplogroup M, dan tempat lahirnya diperkirakan di Papua (dataran rendah). Kekerabatan Indonesia Timur (populasi keturunan Melanesia) dengan Papua Nugini direpresentasikan oleh Q1 dan sedikit Q2, diasumsikan migrasi ke arah barat (NTT, Sulawesi, dan Maluku, serta Filipina) sejak awal Holocene dengan sebaran bahasa Trans-Nugini dan tanaman pisang. mtDNA Q dan P serta Y-DNA S-M30 dan M-P34 migrasi ke arah barat, dibuktikan dengan gen Denisovan yang juga diwarisi oleh populasi Wallacea. Mona et al. (2009) berpendapat bahwa mtDNA Q lahir di Wallacea (berumur 48.089 tahun, ketimbang Papua Nugini (34.678 tahun), meskipun mtDNA Q mendominasi pegunungan selatan PNG dan dataran rendah Papua barat. Keragaman mtDNA Q di Indonesia Timur sekitar 96,7%, lebih tinggi daripada keragaman genetik di Papua Nugini. mtDNA Q dan M29 memiliki nenek moyang yang sama M29’Q yang hanya 1 mutasi dari macrohaplogroup M pada SNP 13500. Q berada 6 mutasi dari M29’Q, sedangkan M29 17 mutasi dari M29’Q. Merriwether et al. mengestimasi umur mtDNA Q 44.500±7400 tahun, sedangkan Hudjashov et al. 32,000±6,500 tahun. Mitogenome dari Behar et al. (2012) mengestimasi umur Q sekitar 37.400 tahun. Friedlaender et al. (2005) mengestimasi mtDNA Q1 49,856 tahun (seumuran dengan P1 51,316 tahun). Umur mtDNA P yang autochthonous di barat daya Pasifik (Papua Nugini dan Papua barat) sesuai bukti arkeologi di area ini (~48-40.000 tahun): berdasarkan coding-region berumur 51.700-65.400 tahun dan berdasarkan mitogenome 54.800 tahun (mendekati umur Friedlaender et al. 2005). Hubungan PNG dengan Indonesia Timur didominasi oleh mtDNA Q dan Q1 serta P dan P1. Beberapa garis keturunan Q dan P di Timor Leste mengkonfirmasi kekerabatannya dengan PNG sudah sejak lama. Wanita Timor Leste dan PNG mengalami isolasi hampir keseluruhan terlihat dalam analisa inti DNA. Hal ini mendukung sejarah populasi berkelanjutan terjadi di Sahul lebih dari 50.000 tahun yang lalu. Gomes et al. (2015) berpendapat bahwa gap yang ada pada garis keturunan maternal P dan Q di Sahul tidak bisa menjelaskan adanya perpisahan nenek moyang mereka yang berpisah sesampainya di Sahul. Karena Gomes et al. tidak mempertimbangkan pendahulu mtDNA P dan Q, yaitu terpisahnya mtDNA M42 (ke selatan) dan M29’Q (terus ke timur dan timur laut) sesampainya di paparan Sahul.
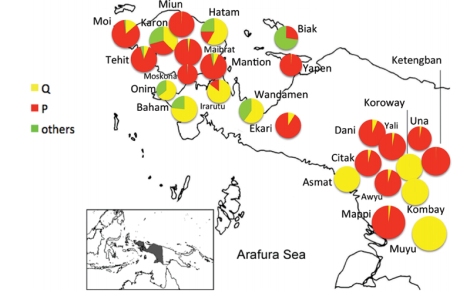
Sebaran mtDNA P dan Q di Papua Barat
Menjelajahi Oceania
Pendudukan Oceania (Melanesia, Polynesia dan Micronesia) sangat terkait erat dengan sejarah populasi Indonesia, karena nenek moyang populasi Oceania pada dasarnya adalah masih garis keturunan populasi gelombang pertama yang menghuni Sundaland, yang juga terkait dengan Orang Asli Semenanjung Malaysia, dan populasi gelombang kedua tersebut, yang kemudian migrasi dari Sundaland ke Sahul, menjadi nenek moyang populasi Oceania. Bisa disimpulkan, bahwa gelombang kedua tersebut adalah penduduk pribumi Sundaland (lihat ulasan Y-DNA K2b1 dan mtDNA P) yang migrasi ke Sahul sekitar 55-50.000 tahun yang lalu.
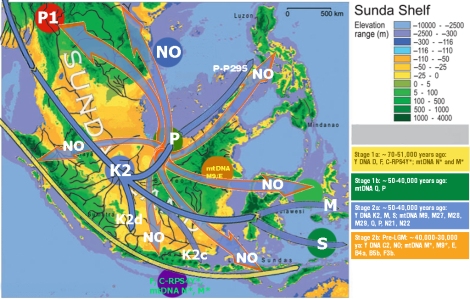
sebaran Y-DNA K2b1
Sebaran mtDNA P di Wallacea dan Sahul
Oceania adalah salah satu wilayah terpenting dalam sejarah sebaran manusia, karena keragaman genetik di wilayah yang meliputi Melanesia dan kepulauan Pasifik, disebabkan oleh setidaknya dua peristiwa migrasi skala besar. Pertama, migrasi dari Sunda ke Sahul oleh populasi pribumi Sundaland yang pada saat ini bisa kita jumpai dalam populasi aborigin Australia, highlander Papua Nugini dan sebagian populasi pribumi Wallacea (Nusa Tenggara, Maluku dan Filipina), dan migrasi kedua, ekspansi penutur bahasa Austronesia dari wilayah Wallacea ke kepulauan Pasifik, dengan jejak sebaran Polynesian motif-nya.
Sebelum 8000 tahun yang lalu, Australia dan Papua masih bersatu sebagai paparan Sahul, dan pada akhirnya menjadi tujuan populasi pribumi Sundaland sekitar 50.000 tahun yang lalu, setidaknya dari bukti arkeologi di Australia (Roberts RG, Jones R, Smith MA, 1990: Thermoluminescence dating of a 50,000-year-old human occupation site in northern Australia), dan sekitar 40.000 yang lalu di Papua Nugini (Groube et al., 1986: A 40,000 year-old human occupation site at Huon Peninsula, Papua New Guinea). Meskipun pendudukan awal Oceania secara rinci, seperti populasi berasal dari mana, jalur-jalur yang mungkin ditempuh, serta kapan migrasi tersebut terjadi, masih diperdebatkan banyak pihak, penelitian berdasarkan penanda genetik maternal (mtDNA) dan penanda genetik paternal (Y-DNA) kadang masih menunjukkan hasil yang bertentangan. Sebagian besar penelitian mendukung pandangan bahwa populasi Australia dan Papua Nugini berasal dari populasi yang berbeda karena tidak adanya kekerabatan genetik di antara mereka (Redd and Stoneking, 1990: Peopling of Sahul: mtDNA variation in aboriginal Australian and Papua New Guinean populations; Huoponen et al., 2001: Mitochondrial DNA variation in an aboriginal Australian population: evidence for genetic isolation and regional differentiation; Ingman and Gyllensten, 2003:Mitochondrial genome variation and evolutionary history of Australian and New Guinean aborigines; Van Holst Pellekaan et al., 2006: Mitochondrial genomics identifies major haplogroup in Aboriginal Australians; Kayser et al., 2001:Independent histories of human Y chromosomes from Melanesia and Australia), sementara penelitian lain – berdasarkan polimorfisme sisipan Alu, dan sebaranHeliobacter pylori – menunjukkan bukti bahwa mereka memiliki nenek moyang yang sama. Perdebatan ini bisa dijawab setelah diperkenalkannya analisisgenome, dengan resolusi yang lebih mendalam dan rinci.
Dua penelitian yang menganalisa data SNP dari populasi aborigin Australia dan Papua Nugini (McEvoy et al., 2010: Whole-genome genetic diversity in a sample of Australians with deep aboriginal ancestry; Pugach et al., 2013:Genome-wide data substantiate Holocene gene flow from India to Australia), meskipun mengkonfirmasi perpisahan populasi pribumi Australia dari populasi Eurasia, mampu mengidentifikasi populasi highlander Papua Nugini sebagai kerabat terdekat populasi aborigin Australia (mtDNA P [Friedlaender et al., 2005], dan Y-DNA K2b1 [Karafet et al., 2014] telah memberikan petunjuk awal bahwa populasi aborigin Australia, highlander Papua Nugini dan Aeta di Filipina berkerabat dekat secara genetik).
Simon Easteal et al juga menemukan diversitas yang ekstrim pada populasi PNG. Mereka mencoba membuat pemetaan garis keturunan di PNG, berhasil mengidentifikasi dua pusat sebaran, haplogroup P dan Q, dengan garis keturunan menyebar ke segala arah. Dua garis keturunan maternal yang nampaknya sangat tua, dan terpisah dari mtDNA di luar Afrika. Menariknya, haplogroup Q tidak ditemui pada garis keturunan maternal di Australia (ada Q2b di timur laut Australia, sepertinya mereka yang mencoba menyeberang ke Australia ketika masih sebagai paparan Sahul, dan berhenti di sekitar Teluk Carpentaria, Queensland bagian utara), dan haplogroup P hanya ditemui beberapa saja, sebagai minoritas di Australia. Hal ini menunjukkan bahwa terjadi isolasi yang cukup ekstrim antara populasi Australia dan PNG walaupun mereka pernah bersatu dalam paparan Sahul.
Pendudukan awal Sahul berdasarkan bukti arkeologi, dan diferensiasi genetik populasi aborigin Australia dan highlander Papua Nugini, membuat para peneliti kemudian mengubah pandangan bahwa sebaran ke arah Pasifik bagian barat (Oceania terdekat) adalah bagian dari migrasi pertama setelah mereka sampai di Sahul (setidaknya 15.000 tahun setelah menghuni Sahul). Pandangan ini tidak memiliki bukti yang cukup, setelah diuji bersama dengan dua hipotesa yang lain, berkenaan dengan asal-usul populasi Papua Nugini, menggunakan sekitar 1 juta SNP dari populasi Oceania (Wollstein et al. 2010: Demographic history of Oceania inferred from genome-wide data). Tiga model diuji, dan model demografik yang didukung analisis SNP menggambarkan perpisahan leluhur Papua Nugini dari leluhur Eurasia; adanya kemungkinan kecil Papua Nugini berkaitan dengan populasi Asia Timur; namun tidak ada bukti bahwa populasi Papua Nugini terkait langsung dengan populasi Afrika.
Analisis genome membantah kemungkinan hipotesis ‘early dispersal’ (oleh populasi Negrito Sundaland berasal dari Afrika), namun untuk mengidentifikasi kemungkinan jalur yang diambil oleh migrasi gelombang pertama juga masih merupakan tantangan yang harus dijawab, sebagaimana bukti arkeologi dari jejak jalur pesisir pantai selatan out of Africa pasti sudah hilang tertelan lautan pada ujung Last Glacial Maximum (LGM), dan sisa-sisa genetik purba bisa jadi telah tergantikan oleh peristiwa-peristiwa migrasi berikutnya.
Berkenaan populasi aborigin Australia dan highlander Papua Nugini, kehadiran kelompok Negrito Malaysia dan Filipina serta pribumi Andaman, diduga adalah keturunan langsung dari diaspora out of Africa melalu jalur pesisir pantai selatan benua Asia, sementara populasi yang hidup di Asia Tenggara saat ini telah terbukti muncul kemudian, sebaran yang berbeda dari arah utara (Lahr and Foley, 1994: Multiple dispersals and modern human origins; Reich et al. 2011: Denisova admixture and the first modern human dispersals into Southeast Asia and Oceania; Rasmussen et al. 2011: An aboriginal Australian genome reveals separate human dispersals into Asia). Kekerabatan genetik antara populasi aborigin Australia dan Negrito Filipina awalnya berdasarkan data Y-DNA (Delfin et al., 2011: The Y-chromosome landscape of the Philippines: extensive heterogeneity and varying genetic affinities of Negrito and non-Negrito groups), dan bukti tersebut diperkuat dengan analisa genome, yang mengungkap afinitas populasi aborigin Australia dan highlanders Papua Nugini dengan populasi Aeta dan Mamanwa, kelompok Negrito dari Filipina, adalah cukup dekat (Pugach I et al., 2013: Genome-wide data substantiate Holocene gene flow from India to Australia; Reich et al., 2011). Untuk menguji kekerabatan genetik antara populasi pribumi Asia Tenggara dan pribumi Eurasia yang datang dari arah utara, perlu kajian tersendiri terhadap mtDNA haplogroup N dan M serta R antara kedua wilayah tersebut, dan menentukan basal garis keturunan yang membedakan kedua garis keturunan itu.
Selanjutnya, data genotip skala besar memungkinkan estimasi waktu perpisahan antara nenek moyang aborigin Australia dengan populasi lainnya. Menggunakan korelasi dalam pola linkage disequilibrium (LD) genome antar populasi untuk memperkirakan waktu divergensi mereka, Pugach et al. mengestimasi populasi Eurasia dan aborigin Australia terpisah secara genetik dari populasi Afrika sekitar 66.000 tahun yang lalu (lebih muda dari perpisahan antara Y-DNA CT dari Y-DNA B dari kajian Karmin et al. 2015 yang menempatkan perpisahan CT dari B sekitar 100.000 tahun yang lalu; perpisahan proto-Eurasia dan pygmy Afrika Tengah), sedangkan perpisahan antara populasi aborigin Australia dan highlanders Papua Nugini dari populasi awal Eurasia sekitar 43.000 tahun yang lalu (terpisahnya Y-DNA Australasia CDF dari Y-DNA Eurasia CEF, serta mtDNA M & N dari L3, menurut Karmin et al. perpisahan terjadi paska letusan Toba 73.000 tahun yang lalu. Sesuai dengan umur mtDNA N dan M sekitar 70.000 tahun yang lalu), dan perpisahan antara populasi Australia, Papua Nugini, dan Mamanwa Negrito diperkirakan terjadi 36.000 (Pugach et al., 2013) (Y-DNA K2b1 dan mtDNA P, menurut Kamin et al. 50.000 tahun yang lalu. Namun menurut Gomes et al. sebelum 37.000 tahun yang lalu). Menurut kajian aDNA dari peneliti Rusia, populasi aborigin Australia menginjak Sahul sekitar 49.200 tahun yang lalu [interval:43.900-54.600 tahun yang lalu]).
Walaupun demikian, umur 43.000 tahun tersebut sesuai dengan estimasi waktu sebaran bakteri H. pylori. Lebih jauh lagi, umur tersebut menunjukkan bahwa populasi aborigin Australia dan highlanders Papua Nugini berpisah setelah mereka sampai di Sahul, ~50.000 tahun yang lalu, dan bukan pada saat permukaan air mulai memisah Sahul 8000 tahun yang lalu. Singkatnya, populasi aborigin Australia sudah sampai di Sahul 43-50.000 tahun yang lalu. Untuk memastikannya, bukti arkeologi sangat dibutuhkan. Manusia purba dari Lake Mungo sebenarnya bisa menjawa teka-teki ini, karena diperkirakan mtDNA manusia purba tersebut lebih tua dari mtDNA N dan M serta berbeda dari garis keturunan L3 dari Afrika yang dipercaya sebagai nenek moyang seluruh mtDNA di luar Afrika.
Menurut bukti arkeologi, kehadiran populasi pribumi di Australasia pada Paleolitik Atas terkonfirmasi dengan temuan di Timor berumur 42.000 tahun, Borneo 46.000 tahun, Papua Nugini 43-49.000 tahun, Melanesia dan Australia lebih dari 48.000 tahun (tersebar di seluruh penjuru Australia). Jadi estimasi Pugach et al. tentang perpisahan antara populasi aborigin Australia, highlanders Papua Nugini dan Negrito Filipina terlalu muda dibandingkan bukti arkeologi.
Dari Sunda ke Sahul
Populasi gelombang kedua ini mendiami Sahul selama ribuan tahun sejak pertama kali menjejakkan kaki mereka (menurut perhitungan berdasarkan aDNA ~49.200 tahun yang lalu), menjelajah setiap sudut Pleistocene Sahul, melahirkan garis keturunan yang hari ini kita kenali dari mtDNA haplogroup P dan subclade-nya tersebar di seantero Australia dan Papua Nugini (Gomes et al., 2015), sampai munculnya penutur Austronesia ~4500 tahun yang lalu, dari sebuah wilayah di Wallacea (Halmahera, bukan Taiwan), yang memiliki dampak cukup besar dari Madagascar di ujung barat sampai pulau Easter di ujung tenggara Pasifik, namun sangat kecil terdeteksi pada populasi Pleistocene Sahul. Mereka hanya terdeteksi di pesisir Papua Nugini, kepulauan Melanesia, dan Polynesia dan kepulauan Pasifik.
Sementara ekspansi Paleolitik di kepulauan Melanesia juga terlahir dari populasi yang berkerabat dekat dengan populasi Pleistocene Sahul (mtDNA Q, M27, M28, M29). Populasi ini terdeteksi sedikit di Australia (hanya Q2b), namun dominan di Papua Nugini, bisa jadi karena garis keturunan tersebut lahir di sana (autochtonous PNG). Mereka kemudian menjadi populasi pribumi Melanesia, sampai datangnya ekspansi Holocene yang memiliki kemampuan dan ketrampilan mengarungi perairan bebas, yang memungkinkan mereka sampai di kepulauan Pasifik, terpisah dari kelompok mereka sendiri, menguasai Pasifik dengan tradisi dan gaya hidup mereka yang dikenal dengan kebudayaan Lapita. Penelitian awal mtDNA dan Y-DNA memberikan bukti bahwa pada saat mereka sampai di Melanesia, penutur bahasa Austronesia tersebut mulai bercampur dengan populasi pribumi penutur rumpun bahasa Papua, dan populasi hasil asimilasi tersebut akhirnya menguasai kepulauan Pasifik (Kayser et al. 2006: Melanesian and Asian origins of Polynesians: mtDNA and Y chromosome gradients across the Pacific; Melton et al., 1985: Polynesian genetic affinities with Southeast Asian populations as identified by mtDNA analysis; Sykes et al., 1995: The origins of the Polynesians: an interpretation from mitochondrial lineage analysis; Redd et al., 1995: Evolutionary history of the COII/tRNALys intergenic 9 base pair deletion in human mitochondrial DNAs from the Pacific; Kayser et al. 2008: The impact of the Austronesian expansion: evidence from mtDNA and Y chromosome diversity in the Admiralty Islands of Melanesia; Kayser et al. 2008: Genome-wide analysis indicates more Asian than Melanesian ancestry of Polynesians; Kimura et al. 2008: Gene flow and natural selection in Oceanic human populations inferred from genome-wide SNP typing; Trent et al., 1998: Globin genes are useful markers to identify genetic similarities between Fijians and Pacific Islanders from Polynesia and Melanesia)
Percampuran populasi yang ekstensif, sebelum ekspansi populasi dengan leluhur dari Asia dan Papua tersebut, tercermin dalam sebuah model asal-usul Polynesia yang dikenal dengan ‘Slow Boat’ (Pellekaan et al., 2006).Namun, populasi campuran ini terbukti tidak sesuai dengan realitas, karena kebanyakan mtDNA di kepulauan Melanesia dan Polynesia saat ini keturunan dari Asia (Wallacea B4a1a1a), sedangkan Y-DNA didominasi keturunan Papua Nugini, dalam hal ini Y-DNA K-M526* dan subclade-nya (M dan S), serta garis keturunan C-RPS4Y*: C-M38 dan C-M208 serta subclade-nya (Kayser et al., 2006, 2008), sesuai dengan pola tradisi matrilokal masyarakat penutur Austronesia. C-M38 dan K-M526* bisa jadi populasi yang sama sejak awal, mempertimbangkan pola mutasi dan diferensiasinya paralel. Keragamannya cukup tinggi di Australasia.
Hal ini menggambarkan migrasi awal populasi yang lebih rumit, diikuti oleh sebaran tunggal dari Wallacea (bukan dari Taiwan) ke kepulauan Melanesia yang menghasilkan percampuran yang ekstensif dengan populasi pribumi sebelum mereka berekspansi ke kepulauan Pasifik. Meskipun demikian, skenario sederhana ini, memberikan kerangka untuk memudahkan memahami warisan genetik migrasi manusia ke Oceania, namun tidak menjelaskan semuanya, sebagaimana bukti arkeologi, bahasa, dan genetik memperlihatkan sejarah populasi yang lebih rumit.
Sebagai contoh, sebaran gerabah khas budaya Lapita yang tersebar luas, terkait dengan ekspansi Austronesia, menambah rumit pola sebaran bahasa Austronesia (Blust 1995: The prehistory of the Austronesian-speaking peoples: a view from language; Bellwood 2004: First farmers: the origins of agricultural societies), dan kehadiran outlier genetik seperti pulau Santa Cruz, di mana mtDNA dan Y-DNA haplogroup dari Papua umum dijumpai di sana, mengindikasikan bahwa skenario dua-gelombang ternyata kurang lengkap. Pulau Santa Cruz, sebagai batas pertama sebelum memasuki kepulauan Pasifik, memiliki nenek moyang yang berasal dari Papua, serta sepertinya bukan merupakan tempat pemberhentian pertama dari para pengembara laut masa lalu tersebut karena mereka melanjutkan penjelajahan di kepuluan Pasifik. Kajian yang lebih mendalam dari variasi regional diperlukan untuk memberikan rincian tentang jalur yang dilalui, kemungkinan adanya migrasi yang lain, serta kontak antar populasi setelah mereka ekspansi ke kepulauan Pasifik.
Data penting tentang asal-usul populasi Polynesia baru-baru ini disimpulkan berdasarkan penelitian 1 juta SNP dari populasi Papua Nugini, Fiji dan tujuh pulau lain di Polynesia, serta satu populasi dari Borneo (Wollstein et al. 2010:Demographic history of Oceania inferred from genome-wide data). Penelitian ini memperkenalkan pendekatan baru untuk menghindari bias. Hasilnya, penelitian tersebut tidak hanya berhasil menghitung persentase percampuran populasi Polynesia yang terdiri dari 85% garis keturunan Asia (Wallacea) dan 15% dari Papua Nugini (dengan populasi Borneo sebagai proxy populasi dengan hasil lebih baik ketimbang Han China), namun juga menunjukkan bahwa setelah pendudukan awal, populasi Fiji mewarisi aliran gen dari populasi kepulauan Melanesia, yang tidak melanjutkan migrasi ke Polynesia, yang juga dibuktikan oleh temuan arkeologi (Kirch 2000: On the road of the winds: an archaeological history of the Pacific Islands before European contact). Estimasi populasi Fiji memiliki garis keturunan dari Polynesia sebesar 63%, dan garis keturunan Papua Nugini sebesar 37%. Estimasi waktu percampuran di Polynesia disimpulkan terjadi sekitar 3000 tahun yang lalu, sedangkan di Fiji terjadi sekitar 500 tahun yang lalu. Kedua waktu tersebut sesuai dengan bukti arkeologi (Kirch 2000). Estimasi waktu untuk Fiji menunjukkan adanya tambahan aliran gen dari Papua Nugini setelah pendudukan awal kepulauan Pasifik.
Meskipun wilayah ekspansi penutur Austronesia sangat luas, mereka tidak meninggalkan jejak di benua Australia. Bahkan sebelum analisis genomediperkenalkan, dipercaya bahwa setelah pendudukan awal benua kanguru tersebut, populasi aborigin Australia mengalami isolasi dari populasi lainnya, sampai kedatangan bangsa Eropa di akhir abad ke-18. Kajian variasi mtDNA (Redd & Stoneking, 1999: Peopling of Sahul: mtDNA variation in aboriginal Australian and Papua New Guinean populations; Kumar et al. 2009:Reconstructing Indian-Australian phylogenetic link) dan Y-DNA (Redd et al. 2002:Gene flow from the Indian subcontinent to Australia: evidence from the Y chromosome) menunjukkan kemungkinan adanya kekerabatan genetik dengan India di era Holocene, namun tidak pernah terbukti sampai data genome populasi aborigin Australia berhasil dipetakan, dan hubungan tersebut bisa diteliti lebih jauh (Pugach et al., 2013). Dengan meneliti populasi Northern Territories, highlanderPapua Nugini, India, serta Asia Tenggara, aliran gen dari India ke Australia bisa dibuktikan dengan 4 model analisis, dan waktu percampuran tersebut diestimasi sekitar 4200 tahun yang lalu. Sejak perdagangan sebelum kedatangan bangsa Eropa mungkin terjadi antara populasi pesisir timur laut Australia dan Indonesia, skenario aliran gen tak langsung melalui kepulauan Asia Tenggara juga dipertimbangkan, namun tidak ditemukan jejak leluhur India pada populasi di kepulauan Asia Tenggara. Menariknya, waktu estimasi percampuran sesuai dengan waktu masuknya dingo (anjing liar) ke Australia, munculnya microliths (alat dari batu ukuran kecil), dan bukti arkeologi lainnya. Mungkin saja bahwa perubahan-perubahan yang terjadi di Australia terkait dengan migrasi dari India, meskipun hal tersebut masih dianggap kontroversial (Brown P, 2013:Palaeoanthropology: of humans, dogs and tiny tools; Price and Bird, 2013:Interpreting the evidence for middle Holocene gene flow from India to Australia;Pugach and Stoneking, 2013: Reply to price and bird: no inconsistency between the date of gene flow from India and the Australian archaeological record)
Contoh sample aborigin Australia yang dianalisa berasal dari bagian barat laut Australia, mungkin menarik untuk diinvestigasi lebih lanjut, untuk mengungkap ada tidaknya hubungan antara India dan Australia. Penelitian lain terhadap populasi pribumi Australia berdasarkan sample dari bagian tenggara Australia (daerah Riverine di New South Wales, Pugach et al., 2013) tidak menemukan jejak genetik dari India, namun hal tersebut bisa jadi karena penelitian tersebut tidak memasukkan populasi dari India dan tidak memiliki data yang bisa dibandingkan. Di sisi lain, analisis genome populasi Australia menemukan indikasi adanya kekerabatan genetik dengan kelompok dari India, namun kesimpulan akhirnya bahwa jejak tersebut merepresentasikan nenek moyang dalam genome populasi Australia tersebut tidak bisa diklasifikasikan ke populasi manapun (Rasmussen et al., 2011).
Analisa genome juga sangat berguna untuk mengungkap struktur populasi yang lebih baik pada populasi Polynesia dan highlander Papua Nugini (Wollstein et al., 2010). Tidak seperti pola struktur populasi yang menggambarkan sejarah peristiwa demografik masa lampau, struktur populasi yang baru ini mampu mengindikasikan adanya praktik-praktik sosial masyarakat seperti perkawinan dalam kelompok-kelompok populasi yang tergabung dalam rumpun bahasa yang sama. Seperti, di Papua Nugini, meskipun mereka dari dua desa berdekatan, mereka terpisah secara kelompok bahasa (Huli vs. Angal-Kewa, keduanya dari cabang Engan dari rumpun bahasa Trans-Nugini). Begitu pula dengan apa yang terjadi di Polynesia, bahwa perpisahan antara populasi Pulau Cook dan yang lain di sekitarnya bisa diketahui.
Berdasarkan data mtDNA dan NRY (Y-DNA, Y-chromosome) (Karafet et al., 2010: Major east-west division underlies Y chromosome stratification across Indonesia, Mona et al., 2009: Genetic admixture history of Eastern Indonesia as revealed by Y-chromosome and mitochondrial DNA analysis) populasi di Indonesia Timur adalah garis keturunan Papua dan Asia. Namun, dan ketika data genomemulai tersedia, menjadi memungkinkan untuk menganalisa pola sebaran garis keturunan Asia dan mengestimasi kapan peristiwa tersebut terjadi, yang pada akhirnya bisa mengungkap yang sebenarnya tentang asal-usul garis keturunan Asia di Indonesia, antara pra-Austronesia vs. Austronesia.
Pola yang muncul dalam analisis adalah bahwa garis keturunan Papua meningkat perlahan (sedangkan garis keturunan Asia menurun) dari arah barat ke timur wilayah Indonesia, dengan proporsi terendah (5.1%) keturunan Papua ditemukan pada populasi Toraja di Sulawesi Selatan (populasi terdekat dengan Wallace’s Line), sedangkan populasi Alor, yang terdekat dengan New Guinea, memperlihatkan proporsi tertinggi (55,4%) keturunan Papua (Xu et al., 2012:Genetic dating indicates that the Asian-Papuan admixture through Eastern Indonesia corresponds to the Austronesian expansion). Pola yang sama ini juga terlihat pada populasi Nusa Tenggara dan Maluku. Dengan analisis genome bisa ditentukan kapan percampuran antara kedua garis keturunan, dan disimpulkan percampuran pertama kali terjadi di bagian barat dari Indonesia Timur (sekitar Alor) sekitar 5000 tahun yang lalu dan kemudian sekitar 3000 tahun yang lalu di kepulauan sekitar Papua Nugini. Hasil tersebut sesuai dengan data bahasa dan arkeologi dengan kedatangan penutur bahasa dan budaya material Austronesia di Indonesia (Diamond J, Bellwood P, 2003: Farmers and their languages: the first expansions, Blust R, 1995: The prehistory of the Austronesian-speaking peoples: a view from language; Gray et al., 2009: Language phylogenies reveal expansion pulses and pauses in Pacific settlement; Bellwood 2004: First farmers: the origins of agricultural societies; Bellwood 1997: Prehistory of the Indo-Malaysian Archipelago) dan menolak pandangan bahwa keturunan dari Asia yang ditemukan di Indonesia Timur tidak ada hubungannya dengan, dan terjadi sebelum, ekspansi Austronesia serta bahwa sebaran bahasa Austronesia bisa dijelaskan hanya dengan pembauran budaya saja. Kecenderungan proporsi keturunan Austronesia dan waktu percampuran menunjukkan bahwa penyebaran petani Austronesia di seluruh wilayah Indonesia terjadi dari barat ke timur. Untuk memudahkan, Anda harus melihat peta genetik Nusa Tenggara, dan Anda akan menjumpai bahwa populasi yang migrasi dari arah barat ke timur adalah garis keturunan Austronesia wanita, yang nantinya akan bertemu dengan garis keturunan pria Papua yang menjadi penduduk lokal di Wallacea.
Informasi penting lainnya adalah analisis laju percampuran pada autosome dan X-chromosome pada populasi Nusa Tenggara dan Maluku. Populasi Nusa Tenggara yang dianalisa adalah kepompok penutur Austronesia, memperlihatkan frekuensi keturunan Asia yang lebih tinggi pada X-chromosome dibandingkan dengan estimasi genome secara keseluruhan, menunjukkan bahwa percampuran dalam kelompok populasi tersebut lebih banyak melibatkan wanita dari Asia (penanda mtDNA penutur Austronesia wanita adalah mtDNA M7c3c, F1a1, B4a1a1, jika mtDNA E termasuk, mereka adalah penutur proto-Malayo-Polynesia atau proto-Austronesia, yang juga diklasifikasikan sebagai mtDNA Asia, walau sebenarnya asalnya dari wilayah antara timur laut Borneo dan barat laut Sulawesi). mtDNA F1a1 juga kandidat untuk penutur Austroasiatik, bersama dengan N9a6 dan R9b.
Pola tersebut tidak terlihat pada populasi Maluku, populasi yang dianalisa adalah penutur bahasa Papua. Perbedaan tersebut, dalam hal percampuran antara kelompok keturunan Papua dan Austronesia, sekali lagi konsisten dengan hipotesis bahwa kelompok Austronesia menganut sistem matrilokal (Jordan et al., 2009: Matrilocal residence is ancestral in Austronesian societies; Hage P & Marck J, 2003: Matrilineality and the Melanesian origin of Polynesian Y chromosomes)
Pentingnya pemetaan migrasi manusia skala besar dalam prasejarah Kepulauan Asia Tenggara digambarkan juga dalam sebuah penelitian yang mendemonstrasikan kesuksesan migrasi di Asia Tenggara sebagai populasi yang garis keturunannya berhasil ditelusuri dari berbagai sumber (Lipson et al. 2014:Reconstructing Austronesian population history in Island Southeast Asia). Dengan metode yang relatif bebas dari ketidakpastian yang berhubungan dengan gender, Lipson berhasil membuat peta populasi campuran, proporsi percampuran, dan asal keturunan dari masing-masing populasi campuran tersebut. Komponen Austronesia, yang terkait dengan populasi aboriginal Taiwan, ditemukan dimana-mana dan terlihat di semua populasi Asia Tenggara (dan Polynesia). Hal ini terjadi karena Lipson mengamati garis keturunan pria. Komponen Papua (Melanesia) terbatas hanya di Indonesia Timur dan Polynesia. Komponen Negrito terlihat dalam berbagai proporsi pada semua populasi di Filipina, dan juga pada populasi Indonesia bagian barat walau dalam proporsi kecil, dan tidak ditemui di Indonesia Timur. Komponen Austroasiatik juga cukup besar di Indonesia bagian barat, namun tak terlihat di tempat lainnya di Kepulauan Asia Tenggara, kecuali pada populasi Manggarai di pulau Flores yang masih dalam wilayah Wallacea. Pada populasi Toraja, agak sulit menentukan apakah garis keturunan pria Negrito atau Papua (Melanesia). Namun jika garis keturunan pria Mandar adalah sebagian Papua, dan tidak ada Negrito, maka tak berlebihan jika Toraja juga memiliki garis keturunan pria dari Papua (C-RPS4Y*, C-M38, F-P14*, K-M526*, M-P34, S-M254).
Penelitian tersebut juga mengestimasi waktu percampuran di Kepulauan Asia Tenggara, namun hasilnya lebih muda daripada estimasi waktu kedatangan penutur Austronesia di Kepulauan Asia Tenggara berdasarkan bukti arkeologi dan bahasa, dan estimasi waktu tersebut lebih muda dari waktu kedatangan penutur Austronesia di Indonesia Timur, Polynesia dan Fiji. Meskipun Lipson et al. berpendapat bahwa estimasi yang lebih muda tersebut mencerminkan aliran gen yang terjadi di masa-masa sejarah yang tidak bisa dideteksi oleh metoda yang lain, namun perlu waspada kemungkinan adanya bias dan keterbatasan metode yang dipakai Lipson et al.
Karena estimasi waktu percampuran yang tidak konklusif, sulit untuk mengungkap pemetaan peristiwa yang menjelaskan perjalanan garis keturunan Austroasiatik di Indonesia bagian barat. Lipson et al. mengusulkan tiga skenario:
- skenario pertama, bahwa ekspansi Austronesia melalui daratan Asia Tenggara, di mana komponen genetik Austroasiatik berasal dan dibawa ke Indonesia bagian barat. Namun, skenario ini tidak menjelaskan ketidakhadiran sinyal Austroasiatik di Indonesia Timur. Dan, jika benar komponen Austroasiatik masuk Indonesia bagian barat bersamaan dengan komponen Austronesia, maka kita akan mendapati proporsi kedua komponen pada populasi keturunannya saling berkorelasi; dan hal tersebut belum terlihat.
- skenario kedua, melibatkan percampuran yang terjadi kemudian di daratan Asia Tenggara, yang tidak bisa dianggap sebelah mata.
- skenario ketiga adalah bawah pada saat terjadi migrasi Austronesia, populasi garis keturunan Austroasiatik sudah tersebar di Indonesia bagian barat, dan memang skenario yang paling kuat, dikarenakan wilayah Indonesia bagian barat masih menyatu dengan daratan Asia Tenggara sebagai Sundaland sampai 8000 tahun yang lalu. Hal ini diperkuat oleh analisis genome akhir-akhir ini, dengan kesimpulan bahwa percampuran pertama (antara penduduk pertama Jawa dengan pendatang Austroasiatik terjadi 650 generasi yang lalu atau sekitar 21.000 tahun yang lalu, dengan asumsi umur generasi dan laju mutasi mengikuti populasi tropis). Dan skenario bahwa populasi keturunan Austroasiatik sudah mendiami Sundaland sejak LGM memang paling kuat, dan menjelaskan proporsi penanda genetik Austroasiatik yang tinggi di Indonesia bagian barat, hampir dua kali proporsi Austronesia, sebagai pendatang baru ~15.000 tahun kemudian.
Dengan demikian, maka populasi proto-Austroasiatik adalah keturunan pribumi Sundaland. Penelitian lebih lanjut tentang korelasi dalam garis keturunan, dan waktu percampuran, harus bisa memastikan asal-usul garis keturunan Austroasiatik di Indonesia bagian barat. Untuk lebih jelas tentang populasi di wilayah tersebut, baca sejarah populasi Indonesia gelombang pertama, atau lebih rincinya dalam penelitian berikut (Hatin et al. 2011: Population genetic structure of Peninsular Malaysia Malay sub-ethnic groups; Metspalu et al. 2011: Shared and unique components of human population structure and genome-wide signals of positive selection in South Asia; Deng et al. 2014: The population genomic landscape of human genetic structure, admixture history and local adaptation in Peninsular Malaysia)
Bersambung Gelombang ketiga (sebaran Austroasiatik)..
Sumber:
https://motherlanders.wordpress.com/2015/04/19/dari-sunda-ke-sahul/
No comments:
Post a Comment