SEJARAH GENETIK ORANG ASLI
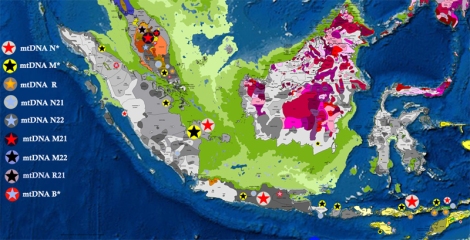
Sebaran mtDNA Orang Asli
Mempelajari sejarah genetik Orang Asli bisa memberikan petunjuk penting tentang pendudukan Sundaland secara keseluruhan. Semua kelompok Orang Aslimengalami pergeseran genetik tingkat tinggi, namun filogeografis mampu menelusuri jejak-jejak garis keturunan maternal dengan cukup meyakinkan.Semang memiliki garis keturunan yang sangat tua di Semenanjung Melayu, yaitu sejak pendudukan populasi pertama 75.000 sampai 50.000 tahun yang lalu. Semang, yang menampilkan fenotipe paling konsisten di areanya, memiliki proporsi garis keturunan pribumi yang tinggi: M21a, M21b, dan R21 (Hill et al., 2006; Oppenheimer 2011). Senoi merupakan gabungan, dengan garis keturunan maternal sekitar setengahnya adalah keturunan Semang dan setengahnya dari Indo China. Hal ini sesuai dengan dugaan bahwa mereka keturunan penutur awal Austroasiatik, yang menyebarkan bahasa dan budaya cocok tanam ke area selatan Semenanjung Melayu sekitar 4000 tahun yang lalu dan bercampur dengan populasi Semang. Populasi aborigin Melayu lebih beragam, berkerabat dekat dengan populasi di kepulauan Nusantara, mereka juga mewarisi haplogroup yang sangat tua dan langka. Kontra dengan ekspektasi sebelumnya, salah satu urutan genom mtDNA mereka, R9b, berasal dari Indo China sekitar Last Glacial Maximum (LGM), diikuti oleh sebaran awal Holocene melalui Semenanjung Melayu ke kepulauan Nusantara.
Walau jumlah Orang Asli hanya 0,5% dari populasi setempat, namun mereka mencakup keanekaragaman fenotipik yang cukup tinggi. Bahasa mereka termasukAslian, cabang dari rumpun Austroasiatik, yang merupakan bahasa seluruh populasi Orang Asli, dan bahasa Melayu yang merupakan rumpun Austronesia. Tradisi Semang diasosiasikan dengan bahasa Aslian Utara, mencari makan di hutan, egaliterisme, patrilinial, and perawakan orang ‘Negrito’. Negrito di Semenanjung Malaysia mempunyai ciri-ciri fisikal yang sangat menyerupai orangPygmy di Afrika dibandingkan dengan kelompok etnik lain di Asia Tenggara. Sebagai tambahan, ciri-ciri mereka juga secara keseluruhannya berbeda berbanding dua kelompok Orang Asli di Semenanjung Malaysia lainnya, yaitu Senoi dan aborigin Melayu. Tradisi Senoi, direpresentasikan oleh Semai dan Temiar, yang dikaitkan dengan bahasa Aslian Tengah, melakukan praktik peladangan berpindah di dataran tinggi, kelompok yang tinggal di rumah panjang, egaliterisme, keturunan kognatik, perawakan bervariasi dari orang Negrito sampai Mongoloid. Tradisi Aborigin Melayu meliputi dialek Melayu (berbeda dari Semelai yang masih memakai bahasa Aslian Selatan), praktik strata sosial, ahli dalam mengumpulkan dan berdagang hasil hutan, melakukan perlawanan keras terhadap Islam dan pendatang yang lain, dan diasosiasikan dengan perawakan Mongoloid.
Teori tentang pendudukan Semenanjung Melayu:
Berdasarkan gaya hidup forager, yang mencari makan di hutan-hutan, serta anatomi postur, Semang dikelompokkan bersama orang Negrito lainnya seperti Aeta di Filipina dan pribumi Andaman, begitu juga dengan orang Melanesia, Tasmania, dan beberapa forager di hutan tropis Australia. Mereka ini diduga populasi gelombang pertama yang mendiami Sundaland sebelum menyebar sampai Pasifik. Gelombang kedua adalah Senoi, yang dipercaya berasal dari India, bersama dengan Veddoid dan forager bertubuh pendek di Asia Selatan, orang Toala di Sulawesi, and sebagian besar suku aborigin Australia. Aborigin Melayu dipercaya adalah migran pertama dari Mongoloid ke Semenanjung Peninsula, sebagai bagian dari pendudukan kepulauan Indo-Malaysia oleh orang berkulit kuning, rambut lurus “proto-Melayu”. Evolusi lanjutan dan ekspansi Deutero-Melayu di Semenanjung Melayu dengan kolonisasi oleh orang Melayu. (Benjamin)
Belwood punya teori lain. Orang Negrito di Asia Tenggara, termasuk Semang, adalah keturunan populasi asli Asia Tenggara yang disebutnya forager “Australo-Melanesia”. Bahasa Austroasiatik dan Austronesia berasal dari China Selatan dan diperkenalkan ke Asia Tenggara pada pertengahan Holocene melalui ekspansi petani Neolitik dengan perawakan Mongoloid. Penutur Austroasiatik melalui jalur daratan ke arah selatan, sementara penutur Austronesia mengambil jalur laut dari Taiwan ke Filipina, dan kemudian ke Indonesia dan Malaysia. Interaksi antara petani pendatang dengan penduduk asli menghasilkan phenotype campuran di beberapa kelompok, yang paling terlihat adalah Senoi, dan juga terjadi pergeseran bahasa pada populasi Semang menjadi penutur Aslian.
Teori lain oleh Rambo (1988) mencoba menjelaskan perbedaan-perbedaan antar kelompok Orang Asli sebagai produk diferensiasi lokal. Semang dan Senoi berasal dari leluhur yang sama, namun dibedakan oleh adaptasi terhadap perbedaan ekologi habitat mereka.
Namun berdasarkan penanda genetik, urutan genome yang ada di Semenanjung Melayu, menunjukkan bahwa sebagian besar adalah keturunan langsung dari garis keturunan maternal dari Eurasia ~60.000 tahun yang lalu (mtDNA M dan N), dan merupakan asli dan unik di Semenanjung Melayu (Macaulay et al. 2005: Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes). Tentu saja Macaulay et al. tidak mempertimbangkan bahwa mtDNA N dan M di Semenanjung Melayu adalah berbeda dari mtDNA N dan M dari Eurasia. Sebagai contoh, M21 dan M22 yang banyak ditemui di Asia Tenggara adalah garis keturunan mtDNA M di daratan Asia Tenggara, bukan dari mtDNA M dari India (Eurasia), sedangkan M31 dan M32 di kepulauan Andaman adalah garis keturunan mtDNA M dari India, bukan Asia Tenggara. Begitu juga dengan mtDNA N dari Eurasia sangat berbeda dengan mtDNA N dari Australasia.
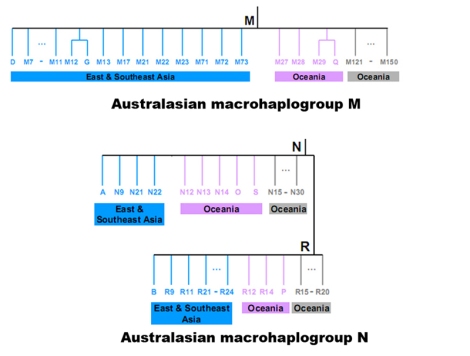
mtDNA M dan N di Australasia dari Kong et al. (2010)
Keragaman garis keturunan macro-haplogroup M
Haplogroup pribumi Orang Asli: M21a (Temuan, Jahai, dan subgroup lainnya di Malaysia Barat. Di luar Malaysia, Sakai dan Chiang Mai di Thailand. Dan agak jarang pada populasi Filipina) dan M22. M21b dan M21c hanya sedikit ditemukan pada populasi Orang Asli, namun banyak ditemukan pada populasi orang laut Moken di Myanmar. M22 ditemukan pada populasi aborigin Melayu, Vietnam, dan China Selatan, belum ditemukan di kepulauan Nusantara.
Haplogroup E, yang menyebar dari perbatasan laut Borneo/Sulawesi ditemukan pada populasi Seletar dan Bidayuh sebagai keturunan, E1b. Garis keturunan haplogroup M7 ditemukan di Malaysia termasuk M7c3c pada populasi Bidayuh. Haplogroup ini hanya ada di Asia Tenggara, dan dianggap sebagai penanda ekspansi penutur Austronesia selama pertengahan Holocene, konsisten dengan umur mtDNA-nya. M7 lainnya pada populasi Temuan M7b1 dan M7c2, dan mereka tergabung dalam garis keturunan populasi dari daratan Asia Tenggara.
Ditemukan juga haplogroup yang tidak pernah ada sebelumnya di Asia Tenggara, yaitu G1c pada populasi Seletar dan M74b serta M20 pada populasi Bidayuh. G1c pernah ditemukan di Korea dan populasi Han di China. Leluhur haplotype M74aada di China Selatan, sedangkan keturunan type M74b ditemukan pada populasi Bidayuh (Serawak, Borneo) dan Hani di China selatan. Sub-type M74b1ditemukan di Surigaono dan Mamanwa di Filipina (dimasukkan dalam M* oleh Gunnarsdottir, 2011) dan juga di Besemah (Sumatra, data Gunnarsdottir 2011 sebagai M4). Dengan ditemukannya cabang paling tua dari M74 di China Selatan, sedangkan subgroup M74b di Bidayuh dan Besemah, menunjukkan bahwa sebaran dimulai dari China Selatan menuju ke arah Sundaland.
Haplogroup M20 di Bidayuh tergabung dengan grup di China Selatan, dan keduanya berkelompok dengan M51 yang ditemukan di Cham (Vietnam) dan Besemah (Sumatra).
Keragaman garis keturunan haplogroup N
Orang Asli: N21, N22 dan R21. Garis keturunan N21 pada populasi Temuan sepertinya keturunan dari haplotype leluhur yang ditemukan di Cham (Vietnam), menunjukkan asal-usulnya dari Indo China pada Pleistocene Akhir berdasarkan estimasi umur DNA. N22 terbatas pada populasi Temuan, sedikit di Filipina, Sumba, dan Sumatra. R21 terbatas pada populasi Negrito di Malaysia Barat, cukup banyak pada populasi Senoi, yang mungkin adalah pendatang dari Indo China. N9a tersebar di Asia Timur, namun subclade-nya N9a6 sepertinya terbatas pada populasi di kepulauan Nusantara (sedikit di Sumatra dan Jawa, namun tidak ada di Filipina dan Taiwan). N9a6 dan keturunannya N9a6a cukup banyak di Malaysia, khususnya Bidayuh dan Seletar.
B sangat umum ditemukan di kepulauan Nusantara, dan Polynesia. Sebarannya bervariasi pada populasi Malaysia, dengan B4a dan B5b di Bidayuh, B4b dan B6di Temuan, dan B4c di Seletar. Dua garis keturunan B4a di Bidayuh termasukB4a1a1a, Polynesian motif, mencerminkan aliran gen dari arah Pasifik (lebih tepatnya Halmahera, Maluku sebagai lokasi munculnya motif) selama periode pertengahan Holocene. Atau, bisa juga populasi in situ garis keturunan mtDNA B4a*. Ada juga B4a yang belum terkelompokkan, berbagi basal mutasi dengan B4a namun tidak diklasifikasikan. Type leluhur dari haplogroup B4b, B4c dan B5b ditemukan pada populasi China Selatan, menunjukkan asal-usul dan sebaran dari daratan Asia Tenggara ke kepulauan Nusantara. Menariknya, B4c2yang ditemukan di Seletar, diekstrak dari rambut kuno populasi Negrito, mengindikasikan terjadinya difusi dari daratan Asia Tenggara pada PleistoceneAkhir.
F sangat umum ditemukan di Asia Tenggara, dengan F1a1a sangat tinggi di Temiar, Senoi. F1a’c berbagi basal mutasi yang sama dengan F1a, kecualinucleotide posisi 4086 yang ada di Bidayuh, Besemah dan Semende.
Yang paling menarik dari penelitian ini adalah terbatasnya keragaman mtDNA populasi Seletar. Hanya ada 4 haplogroup terdeteksi, mencerminkan rendahnya keragaman haplotype, meskipun tidak se-ekstrim populasi orang laut Moken. Keragaman mtDNA yang terbatas di Seletar ini bisa diakibatkan karena pergeseran genetik, ditambah dengan jumlah populasi yang kecil, sekitar 800 individu. Ini bisa menjelaskan kenapa N9a6, yang sedikit ditemukan di kepulauan Asia Tenggara, namun sangat tinggi pada populasi Seletar.
Hasil analysis HVS-I mengindikasikan bahwa mayoritas mtDNA Orang Asli tidak mirip dengan garis keturunan maternal yang ada di Eurasia (bahkan dari Afrika). Bahwa terdapat sejumlah garis keturunan pribumi pada Orang Asli yang bukan dari haplotype asal Afrika, (beberapa menduga dari Eurasia) pada akar haplogroup M, N, dan R. Harus dipahami bahwa Asia Tenggara memiliki akar haplogroup M, N, dan R, yang berbeda dari Eurasia. mtDNA N dari Eurasia tidak akan menurunkan keturunan yang sama dengan mtDNA N dari Asia Tenggara (yang juga disebut dengan mtDNA Australasia).
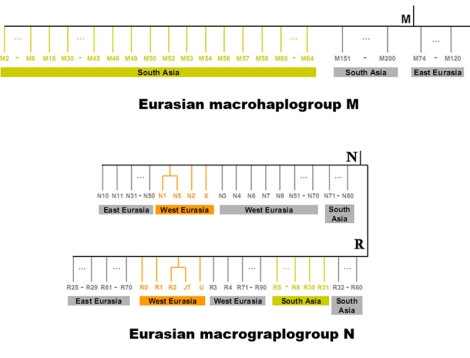
mtDNA macrohaplogroup N dan M di Eurasia dari Kong et al. (2010)
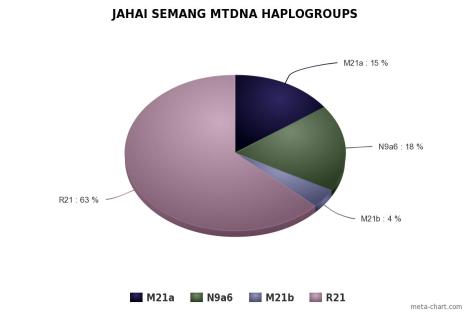
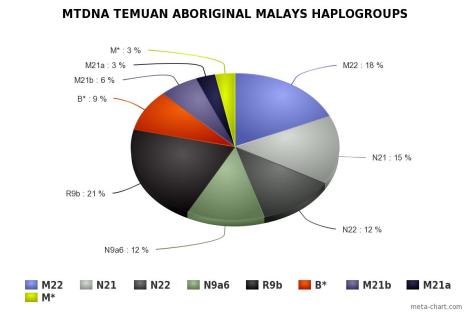
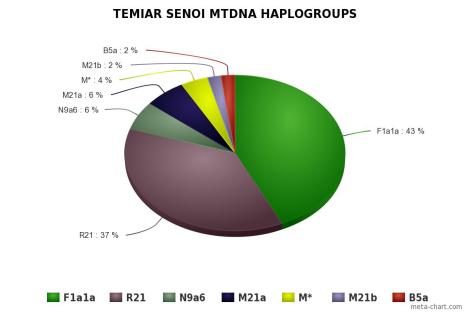
Orang Asli didominasi oleh mtDNA haplogroup M21a pada populasi Mendriq dan Batek Semang, R21 pada populasi Jahai Semang dan Temiar Senoi, F1a pada populasi Temiar Senoi, N21 pada populasi Semelai, M22 pada populasi Temuan, dan R9b pada populasi Aborigin Melayu (Temuan dan Semelai). Dari semuanya, hanya F1a yang berasal dari luar Semenanjung Melayu. F1a berasal dari aborigin Kamboja, sekitar geografi populasi Tompuon (Zhang et al., 2014).
Orang Asli memiliki haplotype yang sama pada tingkat yang cukup tinggi, menunjukkan mereka kehilangan keragaman karena pergeseran genetik, dengan aborigin Melayu lebih beragam daripada Semang dan Senoi. Mendriq memiliki keragaman terendah, dengan jumlah mereka sampai hari ini hanya beberapa ratus individu, di mana >84% peta genetik mereka adalah haplogroup M21a. Yang paling beragam adalah Temuan aborigin Melayu, dan Temiar Senoi berada di antara Mendriq dan Temuan. Perbedaan ini tercermin dalam nilai 3 Orang Aslisecara keseluruhan: keragaman Semang terendah dan keragaman aborigin Melayu tertinggi, dengan Senoi berada di antaranya. Semuanya, secara substansial tidak lebih beragam dari 5 populasi Sumatra (Medan, Pekanbaru, Bangka, Padang, and Palembang). Bisa diinterpretasikan bahwa pergerakan migrasi adalah dari arah Sundaland (lebih tepatnya Sumatra).
Dua haplogroup yang umum dan menyebar luas di Asia Tenggara adalah mtDNA haplogroup B dan R9, dengan R9 mencakup mtDNA haplogroup F.Meskipun teridentifikasi cukup tinggi pada populasi Semai Senoi, haplogroup B hanya ditemukan kecil pada populasi Orang Asli, kecuali B5b yang cukup tinggi pada populasi Batek, mungkin karena pergeseran genetik. Sepertinya B5b masuk ke Batek dari arah selatan (Sumatra, Indonesia), karena hanya sebuah tipe turunan yang hanya ada di satu kelompok etnis, dan sekuens leluhurnya ditemukan di Sumatra dan Indonesia Timur, dan tidak ditemukan di Indo China (Hill, Soares, Mormina, and Richards)
Dua cabang utama haplogroup R9, yaitu R9b dan F lahir sekitar 53.000 tahun yang lalu (Macaulay et al. 2005). Beberapa clade dalam haplogroup tersebut ada pada Orang Asli, masing-masing dengan sebaran yang berbeda. Sebaran R9b jauh lebih luas di Asia Tenggara dibandingkan haplogroup F, sehingga memberikan jalan untuk menentukan waktu pendudukan awal. Di antara Orang Asli, R9b hanya ditemukan pada populasi aborigin Melayu (baik Semelai maupun Temuan) dan sebagian besar diwakili hanya oleh 1 tipe HVS-I, ada pada kedua kelompok. R9b jarang ada di tempat lain tetapi ditemukan dengan frekuensi rendah di Vietnam, Thailand, dan Indonesia (Hill C, Soares P, Mormina M, dan M Richards) dan di provinsi-provinsi Yunnan serta Guangxi di China Selatan.
Munculnya R9b tak lepas dari pra-R9b yang terpisah 29.000 (± 6.600) tahun yang lalu di Indochina dan terpisahnya R9b ~19,000 (± 5.400) tahun lalu di Vietnam / China Selatan. Kemudian satu garis keturunan di mana populasi R9b Thai, aborigin Melayu, dan Indonesia semuanya lahir sekitar 9000 (±2,700) tahun yang lalu. Banyak populasi R9b Indonesia adalah keturunan, dan tidak ada hubungan antara aborigin Melayu dan Indonesia selain mereka memiliki leluhur yang sama. Pola keseluruhan menunjukkan bahwa R9b mengalami diversifikasi di Indo China dan kemudian menyebar ke selatan, Semenanjung Melayu setidaknya 9000 tahun yang lalu, dengan garis keturunannya kemudian menyebar ke seluruh kepulauan Nusantara.
F1a, sangat umum dijumpai dan menyebar di Asia Tenggara, paling banyak pada populasi Senoi, di mana hanya haplogroup F1a1a ada di sana (hampir setengah Temiar dan seorang individu Semai; namun tidak ada pada populasi Semang dan Temuan, meskipun ada jejak kecil di populasi Semelai). Leluhur F1a1a berbagi haplotype dengan populasi Indonesia, Taiwan dan China. F1a1a banyak dijumpai pada populasi Thailand utara dan Vietnam. Keturunan F1a1a ditemukan pada populasi Semelai, dan Senoi. Mereka ada hubungan dengan populasi Nikobar, yang mewarisi F1a1a cukup tinggi. Dengan ditemukannya leluhur mereka di Indo China, hal ini mencerminkan bahwa beberapa populasi Senoi dan Nikobar memiliki leluhur yang sama dari Indo China (lebih tepatnya daerah Kamboja, baca populasi aborigin Kamboja). Leluhur F1a1a lahir sekitar ~10.700 (±4,500) tahun yang lalu, di mana berdasarkan diversitas kontrol-area, umurnya hanya 7000 (±3,000) di Indo China. Hal ini menunjukkan bahwa mereka migrasi ke Semenanjung Melayu dari arah utara (Indo China) dan kawin dengan leluhur Semang, ketika jaman es berakhir.
N9a ditemukan dalam bentuk turunan N9a6a pada 3 populasi Orang Asli dengan jumlah yang sama, meskipun sebarannya tidak sama dan paling beragam pada populasi aborigin Melayu. N9a6a berumur sekitar 5.500 (±2,600) tahun dan sharing dengan populasi Melayu di Malaysia dan Indonesia. Mereka lahir dari N9a6, yang banyak dijumpai pada populasi China Selatan, Indo China, dan Sumatra. Berdasarkan sebarannya, menunjukkan sejarah yang mirip dengan R9b, dengan garis leluhur berada di daratan Asia Tenggara, yang migrasi melalui Malaysia ke kepulauan Nusantara.

Haplogroup N21 lahir tak lama setelah haplogroup N (hanya berjarak 3 mutasi, lihat skema di bawah) ada sekitar 63.000 tahun yang lalu. N21 ditemukan hanya pada populasi aborigin Melayu (Semelai dan Temuan) dan beberapa populasi Melayu di Malaysia (Seletar), dan Indonesia. Meskipun sangat jarang di Indonesia, namun sangat beragam dibandingkan garis keturunan aborigin Melayu, yang juga cukup beragam dalam filogenetik N21, menunjukkan bahwa N21 berasal dari kepulauan Nusantara dan kemudian migrasi ke Semenanjung Melayu. N22 juga ditemukan pada populasi Temuan, di Indonesia jarang namun lebih beragam.

mtDNA Orang Asli
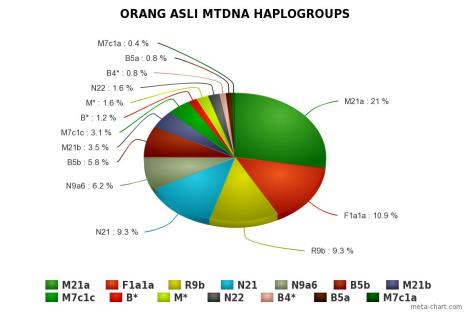
Dalam haplogroup M, terdapat clade lokal yang sangat tua, M21 dengan 3 garis keturunan: M21a, M21b, M21c. M21 berumur sekitar 57.000 tahun yang lalu. M21a paling banyak dijumpai pada populasi Semang (dengan frekuensi cukup tinggi pada populasi Mendriq) dan juga ditemukan pada populasi Mani, populasi Semang di Thailand selatan, menunjukkan bahwa M21a adalah keturunan leluhur populasi Semang. Populasi Mani berbagi tipe sekuens dengan Batek dan Jahai. Pola sebaran sepertinya dari Semang ke Senoi lalu ke aborigin Melayu dan kemudian ke Dayak Borneo Selatan.
M21b lebih jarang, memiliki leluhur yang sama dengan M21a (M21a’b) sekitar 44.000 tahun yang lalu, dan bisa dikatakan haplogroup pribumi Semenanjung Melayu. Dijumpai pada populasi Semang dan Senoi, dengan garis keturunannya ada pada populasi aborigin Melayu dan beberapa populasi kepulauan Indonesia. M21c, saudara M21a’b lebih jarang lagi, ada pada populasi Semelai. Hal ini menunjukkan bahwa bisa jadi adanya hubungan yang cukup tua (pra-glasial) antara kelompok pribumi yang berbeda.
M22 adalah keturunan langsung dari macrohaplogroup M berumur sekitar 63.000 tahun, ditemukan pada populasi Temuan aborigin Melayu dan sedikit di Thai. Haplogroup M yang belum diklasifikasikan dikelompokkan ke dalam M*, dan sebaran filogeografik mereka tidak bisa ditentukan karena tidak adanya tipe urutan HVS-I yang mirip.
R21 kemungkinan besar adalah saudara dekat R9, hanya ada pada populasi Jahai Semang dan Temiar Senoi. R21 lahir dari leluhurnya, haplogroup R, 60.000 tahun yang lalu, meskipun hampir seumuran dengan R9, hanya lebih muda. Seperti M21, sepertinya R21 adalah pribumi Semang/Senoi dan merepresentasikan leluhur yang hidup pada era Pleistocene Akhir di Semenanjung Melayu yang tidak menyebar luas. Ada kemungkinan bahwa mereka hidup terisolasi sejakPleistocene Akhir.
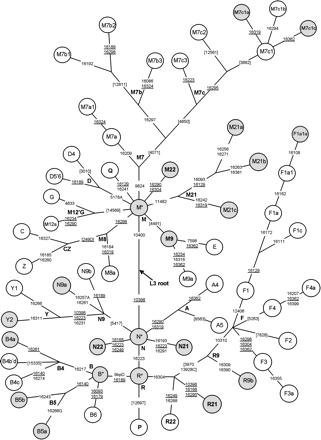
Variasi mtDNA menunjukkan bukti kuat asal-usul Orang Asli di Semenanjung Melayu, sekitar 60.000 tahun yang lalu, tidak harus datang dari Afrika, karena asumsi mutasi balik pada haplogroup N dari L3 harus dibuktikan lebih dahulu. Mari kita asumsikan saja bahwa ada populasi tua di perbatasan China Selatan (koridor Sichuan bagian selatan) dan daratan Asia Tenggara (terutama Thailand) yang sudah ada sejak dulu. Haplogroup M21 dan R21 adalah bukti nyata, yang mendominasi populasi Semang dan Senoi, sedangkan haplogroup N21 dan N22, yang terisolasi pada populasi aborigin Melayu bisa merepresentasikan aliran gen dari kepulauan Nusantara. Aliran gen dari luar: pada semua populasi Orang Asli, terdapat aliran gen era Holocene N9a; Senoi mendapat aliran gen dari Indo China F1a1a; Batek Semang mendapat B5b dari kepulauan Nusantara; dan aborigin Melayu juga mendapat aliran gen dari M7c1c dari arah pesisir, bisa jadi dari ekspansi penutur Austronesia.
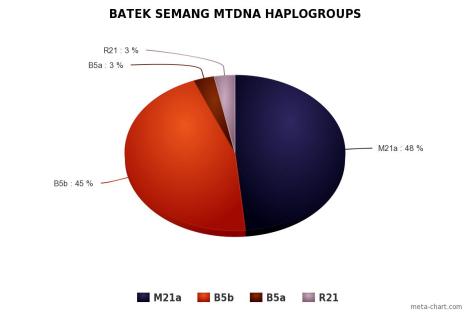
Populasi Semang sepertinya keturunan langsung dari populasi pertama yang mendiami Semenanjung Melayu dan mengalami sedikit aliran gen dari luar. Tiga kelompok etnis Semang berbeda satu sama lain dalam sebaran haplogroup, Jahai misalnya, lebih mirip Temiar daripada Semang. Hubungan mitokondrial populasi Semang sepertinya tidak berkorelasi dengan klasifikasi bahasa, di mana Jahai dan Mendriq adalah satu rumpun bahasa, yang terkait dengan Batek.
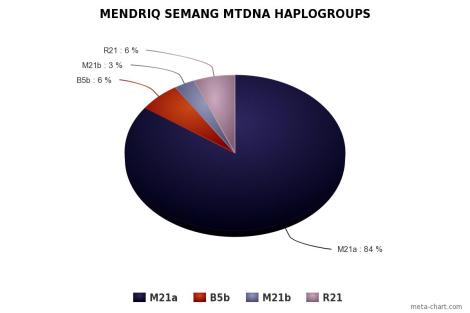
Hal yang penting lagi, tak satupun populasi Semang mirip dengan populasi pribumi Andaman yang juga memiliki haplogroup M pribumi, M31 dan M32. Berdasarkan penanda genetik populasi Aeta Filipina, data genetik kurang sependapat dengan anggapan bahwa populasi Negrito di Asia Tenggara (Andaman, Semenanjung Melayu dan Filipina) memiliki leluhur yang sama. Populasi Andaman dan Filipina bisa jadi satu garis keturunan macrohaplogroup M dari India karena adanya beberapa haplogroup yang satu leluhur, seperti M80’D: M80 ada di Palawan, dan D ada di India Timur. M19’53: M19 di Palawan, dan M53 di Orissa serta India Tengah. M24’41: M24 di Palawan, dan M41 di Bihar, Orissa, Andhra Pradesh serta India India. M31: M31a1 di Andaman, dan M31b’c di India Timur dan India Timur Laut, M31b di Tibet dan Nepal, serta M31a2 di Orissa. M32’56: M32a di Andaman, M56 ada walau minoritas di Indiai Tengah.
Signal demografik yang berbeda sepertinya mengindikasikan bahwa sebaran haplogroup R9b cukup tinggi ditemukan pada populasi aborigin Melayu, dan mungkin juga N9a. Dari sekuens-nya, R9b berasal dari Indo China di eraPleistocene, dengan sebaran ke arah selatan pada awal Holocene melalui Semenanjung Melayu dan kemudian ke kepulauan Nusantara. Pola sebaran ini bertentangan dengan pandangan bahwa populasi aborigin Melayu datang dari kepulauan Nusantara sejak pertengahan Holocene, sebagai salah satu akibat dari ekspansi penutur Austronesia dari Nusantara (Bellwood). Dukungan dari perspektif arkeologi, Hoabinhian berasal dari China Selatan sebelum menyebar ke Melayu dan Sumatra Utara (pada era Pleistocene Akhir/Holocene Awal). Di sisi lain, N21, N22 dan M7c1c berasal dari luar Semenanjung Melayu, sekitar pertengahan atau akhir Holocene, dalam konteks populasi aborigin Melayu.
Signal paling mencolok sepertinya kehadiran F1a1a, di samping pribumi R21, adalah haplogroup paling banyak pada populasi Senoi (hampir setengahnya). Haplogroup ini umurnya sekitar pertengahan Holocene, juga ditemukan cukup tinggi di Indo China dan menyebar di sana dari China Selatan (yang jarang namun lebih beragam, dan di mana leluhurnya ditemukan) selama era Holocene. Hampir setengah dari garis keturunan maternal Senoi bisa ditelusuri kembali asal-usulnya di Indo China sekitar 7000 tahun yang lalu. Hal ini konsisten dengan pandangan Bellwood bahwa budaya Neolitikum dibawa ke Semenanjung Melayu dari Thailand tengah (terkait dengan budaya Neolitik Ban Kao), yang kemudian kawin dengan kelompok pribumi untuk kemudian melahirkan leluhur populasi Senoi sekarang. Populasi ini bisa jadi juga yang menyebarkan bahasa Austroasiatik di Semenanjung Melayu.
Harus diingat bahwa ketiga Orang Asli kemungkinan mengalami pergeseran genetik, sebagaimana diindikasikan oleh pola keragaman mtDNA dan data osteologi. Orang Asli yang semi-terisolasi dan jumlahnya sangat kecil juga ditunjukkan oleh data etnografi. Senoi dan aborigin Melayu tidak mengalami pergeseran yang ekstrim seperti Semang. Namun Senoi mengalami pergeseran genetik lebih dari aborigin Melayu. Hal ini bisa jadi karena proses awal etnogenesis, atau ekspansi Temiar ke arah timur.
Analisis filogeografis menunjukkan setidaknya ada 5 kali peristiwa pendudukan yang mempengaruhi Orang Asli, lebih dari 50.000 tahun yang lalu, pada jaman es terakhir (LGM), sekitar 10.000 tahun yang lalu, pertengahan Holocene, dan akhir jaman Holocene. Ketiga Orang Asli memiliki leluhur lokal setidaknya berumur 50.000 tahun, dan semuanya terdampak oleh migrasi yang terjadi di Semenanjung Melayu setelah itu, besar maupun kecil. Perbedaan-perbedaan di antara Orang Asli mencerminkan leluhur yang berbeda dibandingkan model etnogenesis lokal. Model Bellwood terlalu sederhana; tidak mendeteksi adanya migrasi dari arah utara, yang diasosiasikan dengan budaya Hoabinhian, ribuan tahun sebelum migrasi Neolitik yang disarankan Bellwood. Penting juga untuk mengetahui evolusi lokal dari Orang Asli, dari setidaknya awal Holocene sampai sekarang, yang berpengaruh terhadap gen populasi Senoi dan aborigin Melayu, dan dari kepulauan Nusantara, yang khusus berpengaruh terhadap aborigin Melayu.
Jelas sekarang, bahwa Orang Asli merepresentasikan proses demografi yang terjadi di Asia Tenggara (khususnya prasejarah Sundaland): beberapa garis keturunan maternal yang bisa ditelusuri kembali ke jaman pendudukan awal lebih dari 50.000 tahun yang lalu; beberapa merepresentasikan persebaran pada glasial akhir dan awal Holocene; dan beberapa menunjukkan pergeseran populasi di jaman Neolitik atau paska-Neolitik, dan mungkin juga penyebaran bahasa. Bukti migrasi 10,000 tahun yang lalu di Semenajung Melayu membuka pikiran, memberikan pandangan baru dalam menginterpretasikan kebudayaan Hoabinhian, kemungkinan sebaran forager Asia Tenggara beradaptasi dengan bentang area vegetasi yang berbeda sebelum naiknya permukaan laut di awal Holocene, dan variabilitas osteologis dari ditunjukkan oleh sisa-sisa manusia di akhir Pleistocenedan awal Holocene Asia Tenggara.
Sumber: https://motherlanders.wordpress.com/2015/03/25/sejarah-genetik-orang-asli/
No comments:
Post a Comment